当前位置:网站首页>Nature Microbiology | 可感染阿斯加德古菌的六种深海沉积物中的病毒基因组
Nature Microbiology | 可感染阿斯加德古菌的六种深海沉积物中的病毒基因组
2022-07-04 16:46:00 【刘永鑫Adam】

翻译:周之超@UW-Madison
Genomes of six viruses that infect Asgard archaea from deep-sea sediments
DOI:10.1038/s41564-022-01150-8
原文链接:https://doi.org/10.1038/s41564-022-01150-8
第一作者:Ian M. Rambo
通讯作者:Brett J. Baker
主要单位:Department of Marine Science, University of Texas Austin(德克萨斯大学奥斯汀分校海洋科学系)
摘要
阿斯加德古菌是全球分布的与真核生物有关的原核微生物;然而,感染这些生物的病毒还没有被描述。在这里,利用从深海热液沉积物中提取的宏基因组序列,我们描述了6个相对较大(达117 kb)的双链DNA(dsDNA)病毒基因组,它们感染了两个阿斯加德古菌门,Lokiarchaeota和Helarchaeota。这些病毒编码类似于Caudovirales的结构蛋白,以及与已知古生物病毒中描述的蛋白不同的蛋白。它们的基因组包含与真核细胞质大DNA病毒(NCLDVs)相关的大约1-5%的基因,似乎能够进行半自主的基因组复制、修复、表观遗传修饰和转录调控。此外,Helarchaeota病毒可能劫持宿主的泛素系统,与真核生物病毒类似。对这些阿斯加德病毒的基因组分析显示,它们同时含有原核和真核病毒的特征,此次研究提供了对其潜在感染和宿主互动机制的新见解。
正文
阿斯加德古菌是全球分布的微生物,与真核生物有关。它们的基因组组成表明它们是产生第一个真核生物共同祖先的古细菌宿主的后代。近年来,由于从一系列海洋和陆地水生沉积物中恢复了基因组,阿斯加德的生物多样性大大扩展。一个厌氧的、生长缓慢的阿斯加德谱系,Lokiarchaeota,最近被培养出来,似乎与细菌有互养的依赖关系。这支持了多种基于基因组的阿斯加德-细菌相互作用的推论,这些推论被认为导致了第一个含有线粒体的真核细胞的形成。还有人假设,与病毒的相互作用促成了复杂细胞生命的起源。这是基于一些NCLDVs和噬菌体的核状病毒工厂的存在,它允许在宿主细胞质内复制,并通过mRNA加帽使转录和翻译脱钩。在Lokiarchaeota的基因组内已经发现了推测的病毒蛋白,这表明病毒在遗传元素的交换和阿斯加德古菌的进化中发挥了作用。I型和III型CRISPR-Cas免疫系统已在几个阿斯加德门(Odinarchaeota、Thorarchaeota、Lokiarchaeota和Helarchaeota)中被描述,但迄今尚未对与阿斯加德古菌相关的病毒基因组进行表征。
为了探索病毒在阿斯加德古菌中的作用,我们搜索了Guaymas盆地(~2,000米水深,加利福尼亚湾)热液相关沉积物中~5TB的宏基因组序列数据(assemblies)。从中,我们恢复了6,756个未培养的病毒基因组(UViGs)(图1),估计是高质量和中等质量的(见方法)。UViGs与来自Guaymas盆地宏基因组组装基因组(MAGs)的CRISPR间隔序列(spacer)相连,以确定哪些病毒感染了阿斯加德古菌细胞。这揭示了7个双链DNA(dsDNA)UViG,它们感染了Lokiarchaeota、Helarchaeota和Thorarchaeota(图2)。有趣的是,两个与阿斯加德古菌有关的病毒也感染了从Guaymas盆地重建的Chloroflexi和Omnitrophica细菌,表明阿斯加德与这些细菌有密切联系。其中一个UViG是Chloroflexi基因组中的整合型原病毒,与Thorarchaeota有CRISPR间隔序列联系,而其他六个是非整合型的,被归类为裂解型。我们暂且以北欧神话中的生物命名这些UViG。“Vedfolnir'”(Chloroflexi provirus与Thorarchaeota相关联)、Fenrir(Lokiarchaeota)、Sköll(Lokiarchaeota)、Nidhogg(Helarchaeota)以及Ratatoskr(Helarchaeota和Omnitrophica)。它们的基因组大小(约21.9-117.5kb)属于已知的古菌dsDNA病毒的范围(图2)。从Meg22_1012号样本中发现的Nidhogg和Fenrir病毒与Meg22_1214号样本的核苷酸一致性很高(分别为99.98和99.90%)。Nidhogg病毒被封闭成完整的环形基因组(图2D),两个线性的Fenrir病毒被预测为>90%的完整性。
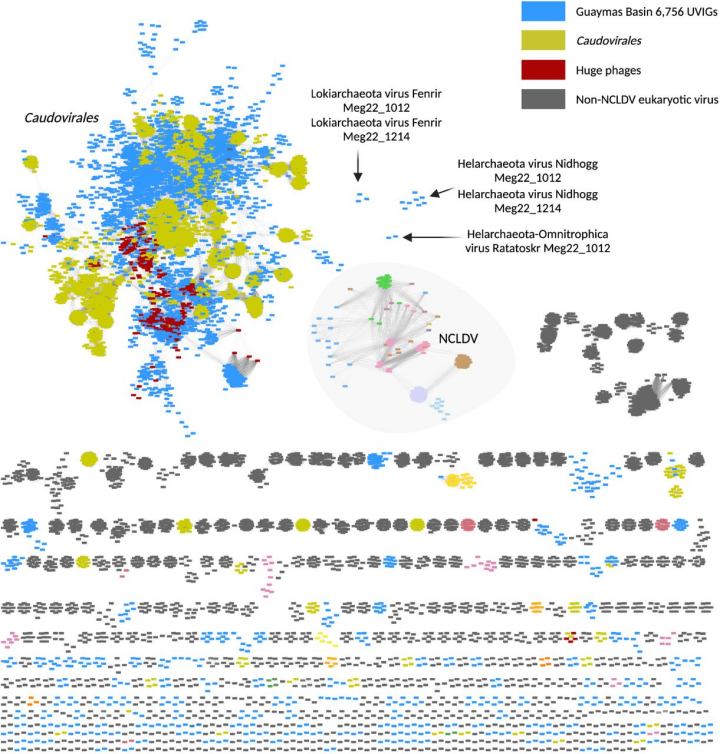
图1. 从Guaymas盆地沉积物中发现的病毒基因组与以前描述的病毒相比的蛋白质聚类网络。
用vContact2 v.0.9.19和Cytoscape v.3.8.0构建的单体网络分析,对Guaymas盆地的UViG(n = 6,756,以浅蓝色显示)进行分类分配,使用来自RefSeq的参考真核生物、细菌和古菌病毒基因组(n = 11,082)和巨大噬菌体(n = 361)(见方法)。节点代表单个基因组,边缘表示病毒簇内基因组之间的相似性(n = 770)。与阿斯加德古菌有关的病毒被标记为Fenrir、Nidhogg和Ratatoskr。Vedfolnir和Sköll被列为离群值(outliers)。
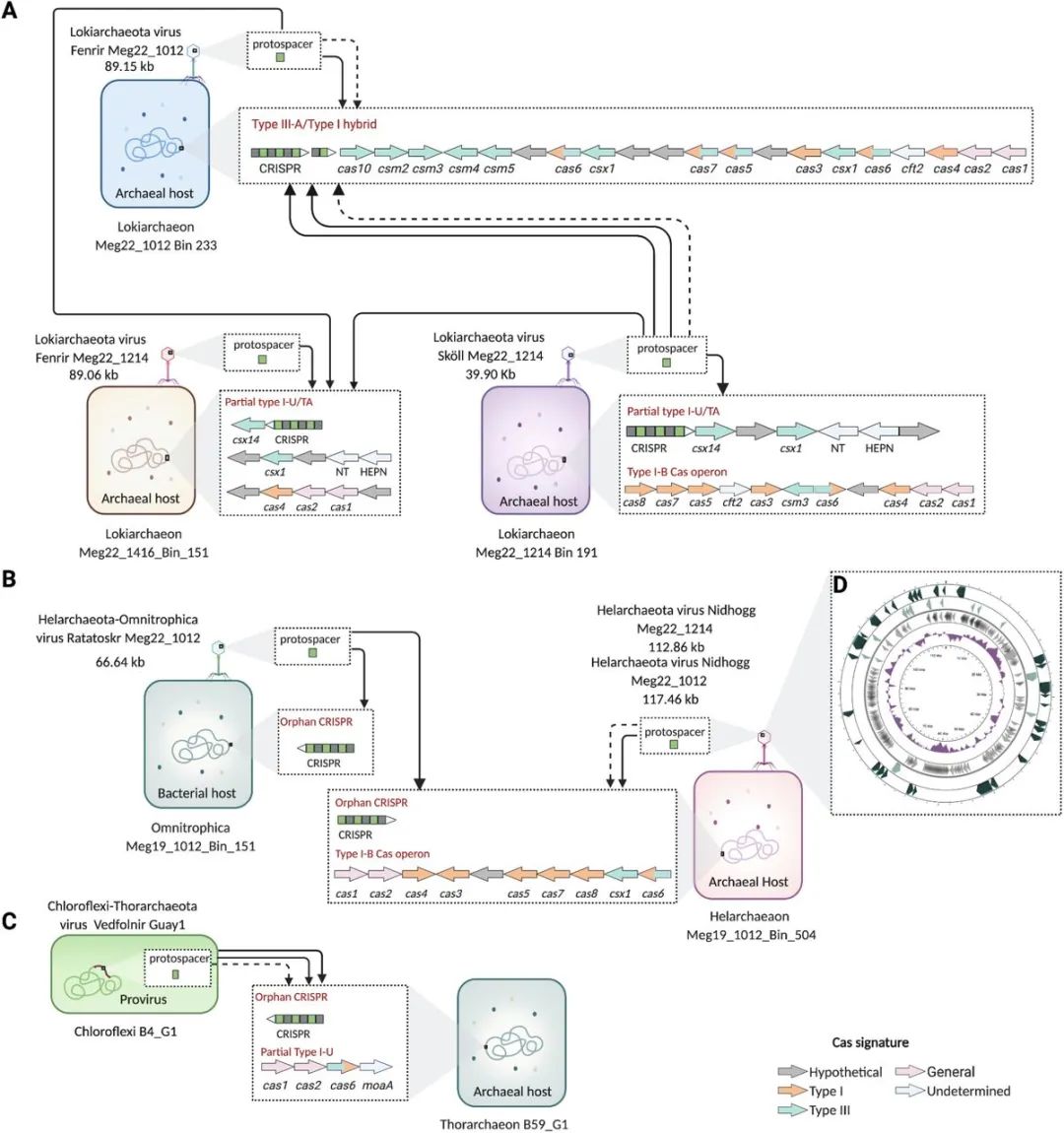
图2. 病毒-宿主与阿斯加德古菌的联系。
(A)Lokiarchaeota, (B)Helarchaeota, 和(C)Thorarchaeota MAGs通过CRISPR-Cas系统与UViG相连。(D)完整的环状Nidhogg病毒基因组的概述。CRISPR间隔条与UViG的blastn-short alignments显示,箭头代表100%的一致性(实线)和95%-99.9%的一致性(虚线)。本研究中回收了Lokiarchaeaota、Helarchaeota和Omnitrophica MAGs。Chloroflexi和Thorarchaeota的基因组先前已被恢复。预测的阿斯加德MAGs的CRISPR-Cas系统由箭头表示,根据其Cas基因的类型特征着色,箭头指向右侧为(+),即正义,左侧为(-)即反义。CRISPR阵列末端的箭头以同样的方式表示正义或反义。显示的每个cas盒都位于不同的scaffold上。病毒用类似噬菌体的图标表示,尽管只有Ratatoskr包含一个可识别的二十面体的外壳。Nidhogg基因组内的编码区域指向右侧为(+),左侧为(-),并按比例绘制。cas1,CRISPR相关内切酶Cas1;cas2,CRISPR相关内切酶Cas2;cas3,CRISPR相关内切酶/螺旋酶Cas3;cas4,CRISPR相关外切酶Cas4;cas5,CRISPR系统Cascade亚单位Cas5。cas6,CRISPR相关内切酶Cas6;cas7,CRISPR-Cas第一类效应器复合体亚单位Cas7;cas8,CRISPR相关蛋白Cas8;csm2,第三类CSM效应器复合体小亚单位Csm2;csm3,第三类RAMP超家族CSM效应器复合体Csm3。csm4, III型RAMP超家族CSM效应器复合物Csm4; csm5, III型RAMP超家族CSM效应器复合物Csm5; csx1, CRISPR系统内切核酸酶Csx1; csx14, III-U亚型相关蛋白Csx14。cft2,Cft2家族RNA处理外切酶;MoaA,钼辅助因子生物合成蛋白MoaA;TA,毒素-抗毒素;NT,核苷酸转移酶;HEPN,高等真核生物和原核生物核苷酸结合域;kb,千碱基。用BioRender.com创建。
为了分类和了解阿斯加德病毒的进化历史,我们确定了七个UViG中的DNA聚合酶B(PolB)蛋白。来自Fenrir、Nidhogg和Ratatoskr的PolB蛋白序列与来自细胞生物、噬菌体、古菌和真核生物病毒以及NCLDVs的PolB进行了对比。这些序列的系统发育分析显示,Fenrir、Nidhogg和Ratatoskr与以前的特征病毒不同,具有复杂的进化历史(图3A)。Fenrir PolB与Gammaproteobacteria的密切关系表明,这些基因是由细菌水平转移而来的,而Nidhogg PolB似乎与宿主为盐杆菌的myoviruses有系统发育关系(图3A)。Ratatoskr PolB在系统发育上与Heimdallarchaeota有关,Heimdallarchaeota是被认为与真核生物关系最密切的Asgard门(图3B)。为了确定阿斯加德病毒的科水平分类,我们对其病毒蛋白家族(VPF)的组成进行了表征。所有七种Asgard UViGs都含有与Myoviridae、Siphoviridae和Podoviridae以及与NCLDVs和嗜热古菌病毒科水平相关的基因同源物(图3C)。Asgard UViGs不与基于单方基因共享网络的现有参考病毒聚类(图1),这可能表明它们是Caudovirales的新科或属。这一点得到了以往研究的支持,这些研究确定了全球性的Caudovirales和古菌之间的关联。与以前描述的病毒缺乏明确的隶属关系,突出了这些新发现的阿斯加德病毒的新颖性。
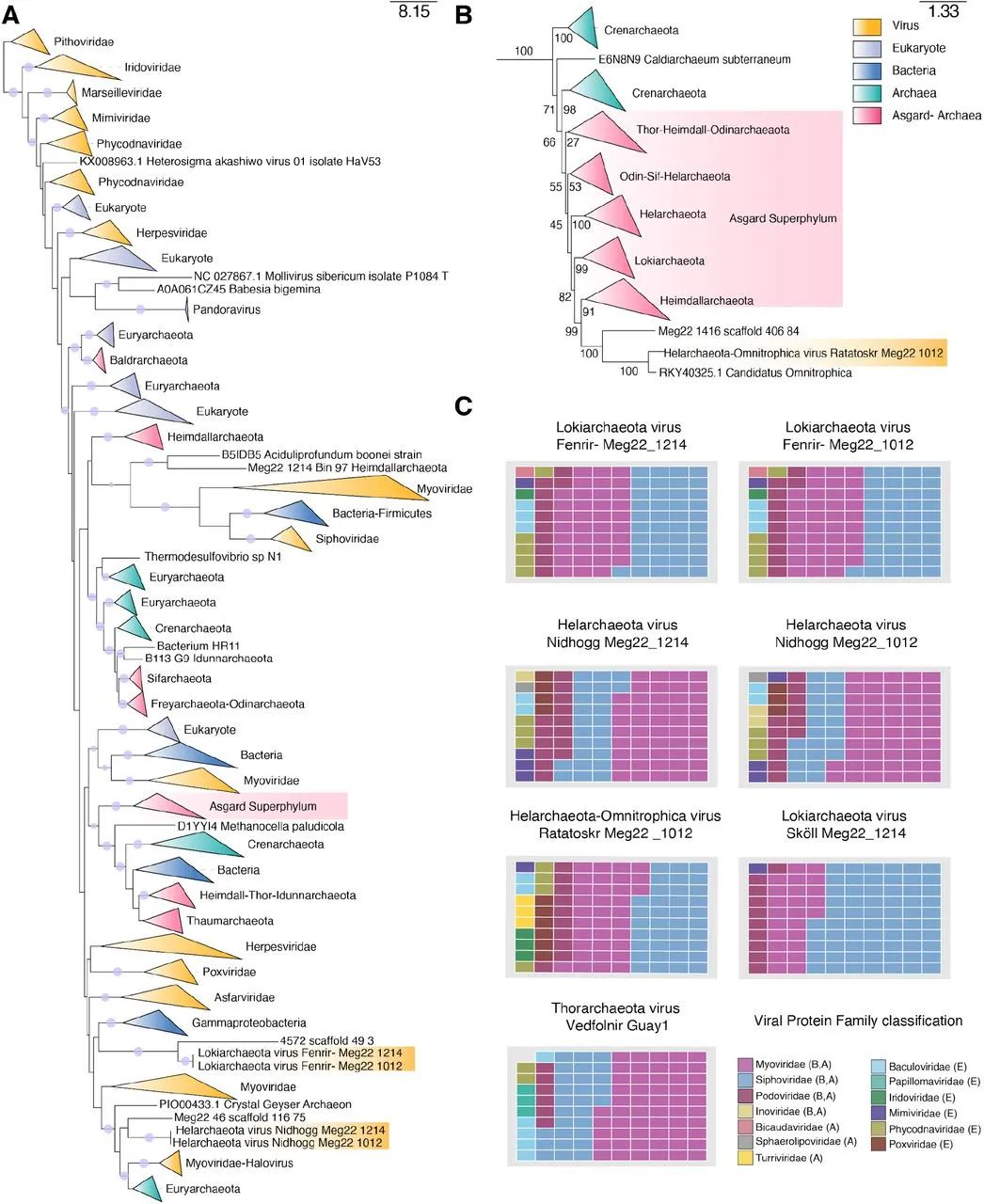
图3. 基于系统发育和蛋白质组成的阿斯加德病毒的分类位置。
(A) 使用LG+F+R10模型生成的DNA聚合酶B的最大似然系统发育树。树枝上的圆圈代表ultrafast bootstrap支持率>=95。本文中描述的病毒用金色突出显示。(B) Ratatoskr在A部分所示的LG+F+R10模型树中的位置,Bootstraps以树枝上的数值显示。(C) 阿斯加德病毒中共享的VPF的成员比率。病毒蛋白家族分类图例中的A、B或E表示古菌、细菌和/或真核生物宿主。
我们在Asgard UViGs中发现了Caudovirales的标志蛋白,包括小头蛋白、基底板J、尾部纤维、门户蛋白和终止酶大亚基(图4)。在Ratatoskr、Nidhogg和Vedfolnir中发现了编码病毒蛋白外壳的推测的衣壳蛋白。在Ratatoskr中发现了一个HK-97-折叠的主要衣壳蛋白(MCP)。在Nidhogg和Vedfolnir(PhANNs,分别为80%和90%的置信度)中检测到的推测的MCPs与收缩性噬菌体尾鞘样蛋白(>99.9%的概率,e-值<4.4e-24)和噬菌体原壳蛋白酶(99.5%的概率,e-值1.6e-13)有结构相似性(HHPred,数据S5)。Fenrir和Ratatoskr都编码HNH内切酶,在包装过程中可能将DNA裂解成基因组长度的单位,并可能与它们的终止酶大亚基和门户蛋白协同运作。然而,在Fenrir和Sköll的基因组中没有发现推定的MCPs,这可能是由于在古菌特异性病毒中普遍存在着新的病毒结构。MCPs的缺失可能是由于含衣壳的病毒和无衣壳的自私元素(capsidless selfish elements,CSE)的紧密进化联系。含衣壳的古菌病毒通过衣壳基因的获得和丧失,经历了向CSE的多次来回转变,就像在真核病毒中看到的那样,但仍然寄生在其宿主的遗传信息中。
为了了解阿斯加德病毒如何影响它们的宿主,我们搜索了存在于原核生物病毒和NCLDVs中涉及基因组复制、核苷酸代谢、转录和宿主-病毒相互作用的基因(图4)。阿斯加德病毒编码原核和真核病毒中存在的核心DNA复制基因(PolB、古/真核引物酶、钳形装载器、RNase H和ATP依赖的DNA连接酶)(26)。据预测,Nidhogg病毒有参与自主基因组复制和校对的基因,包括一个具有转录因子S-II类锌指结构域的ATP依赖性DNA连接酶(ligD)、RNase HI/逆转录酶样蛋白、PolB和引物聚合酶(PrimPol)。据预测,病毒的PrimPol样蛋白在DNA和/或RNA引物活动中具有额外的作用,以及耐破坏的DNA聚合酶活性。迄今为止,这些基因还没有在古菌病毒中被描述过,只在棒状杆菌病毒BFK20和Acanthamoeba polyphaga (棘变形虫)mimivirus病毒中被鉴定过。Ratatoskr UViG编码一个古菌-真核生物DNA引物酶,用于DNA复制中的引物活动,同时也编码PolB。Fenrir和Nidhogg编码AAA ATP酶,与复制因子C小亚基(rfcS)和大亚基(rfcL)相似,可能具有钳形装载器的功能。Sköll有增殖细胞核抗原(PCNA),与来自深海沉积物的Lokiarchaeota关系最密切(蛋白质相似度为36.5%)。PCNA是真核生物和古菌中一个重要的复制过程因子,并存在于它们的一些病毒中。Sköll还编码一个与核糖核苷还原酶(核糖核苷二磷酸还原酶,β亚基)和hedgehog/intein超家族结构域相似的蛋白质。核糖核苷还原酶是由一个保守的核心基因编码,用于NCLDVs的核苷酸代谢。在这个核糖核苷酸还原酶中存在一个hedgehog/intein(Hint)蛋白结构域,可能表明这个基因是从以前的宿主转移到Sköll的。Fenrir病毒在复制过程中可能通过一个类似于细胞膜5'(3')脱氧核糖核酸酶的卤素脱卤酶(HAD)超家族蛋白来调节dNTP池。Fenrir还含有一种脱氧核苷单磷酸激酶(DNMP激酶),与一种大湖藻病毒脱氧核苷单磷酸激酶(bitscore=115.9)和几种Mimiviridae DNMP激酶相似。Fenrir、Nidhogg和Ratatoskr病毒拥有DNA修复基因,这是NCLDVs的一个主要特征。这与缺乏这些基因的小型病毒形成对比。Fenrir、Nidhogg和Ratatoskr病毒还编码ERCC4内切酶域蛋白,与非洲猪瘟内切酶EP364R相似。Nidhogg和Fenrir编码的ERCC4内切酶与古菌XPF 3'瓣修复内切酶相似。这些类似XPF的内切酶含有螺旋-发夹-螺旋结构域,能够实现非特异性DNA结合。
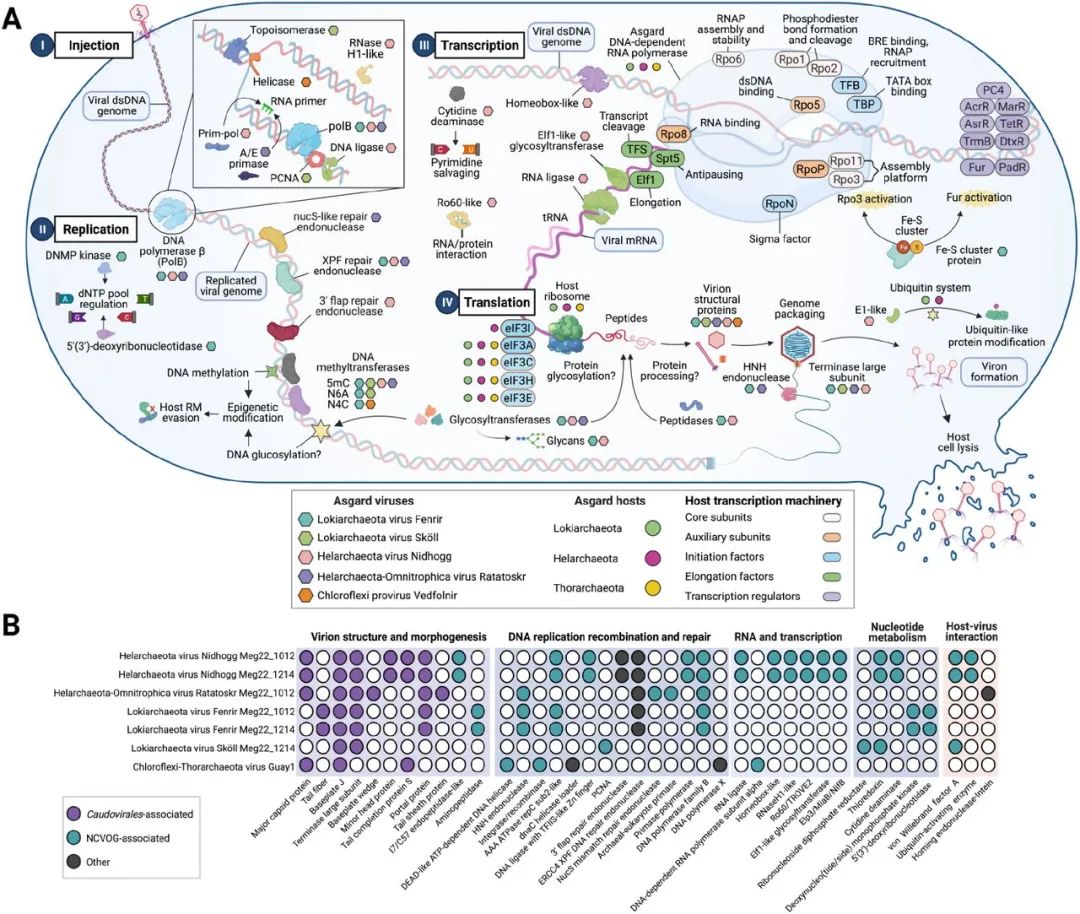
图4. 阿斯加德病毒感染机制的模型以及与其他病毒的蛋白质相似性。
(A)阿斯加德病毒中确定的参与感染、复制、转录和翻译的基因。阿斯加德病毒显示为不同颜色的六边形,阿斯加德病毒宿主显示为不同颜色的圆圈,宿主的转录机器用按功能编码的椭圆来标记(见图例)。(B)40个基因(X轴下部)在阿斯加德病毒(Y轴)中的分布。填充的圆圈表示有注释的直系同源物,而白色的圆圈表示没有发现直系同源物。缩写:NCVOG,核质巨DNA病毒蛋白质同源物群。用BioRender.com创建。
除了核心的DNA复制机器和修复基因外,Fenrir和Nidhogg病毒还有各种基因表达和翻译后修饰的机制。Nidhogg可能通过一个homeodomain蛋白调节转录。Homeodomain蛋白是DNA结合的转录因子,存在于真核生物和Mimiviridae中。Nidhogg病毒编码一个具有TROVE和von Willebrand因子A结构域的Ro60蛋白。Ro60可能与非编码Y RNA形成核糖核蛋白复合物,与蛋白质和其他RNA相互作用。Nidhogg病毒还可能通过RNA连接酶修复、拼接和编辑转录本(图4)。Fenrir编码一个翼状螺旋-转折螺旋结构蛋白,这是在古菌病毒转录因子中发现的一个常见的DNA结合motif。我们没有在这些UViG中发现任何翻译机器,这表明它们利用宿主核糖体和阿斯加德宿主中存在的真核生物启动因子。然而,Nidhogg和Fenrir可能用编码的肽酶处理病毒蛋白前体。据预测,Nidhogg编码的肽酶与Vaccinia病毒(NCLDV病毒的一个种)I7L/C57家族的加工内肽酶相似,而Fenrir似乎能够产生M42家族的氨基肽酶(图4)。
几个阿斯加德门编码真核生物泛素蛋白修饰系统的同源物。真核生物病毒已被证明可以劫持这些系统来帮助病毒传播的各个阶段。Nidhogg编码三个泛素激活(ThiF家族/E1-like)蛋白同源物,可以利用Helarchaeota泛素系统(E1-like激活酶、E2-like连接酶和泛素类蛋白结构域),并将其重新用于复制、转录、病毒子形成和释放等过程。对Nidhogg蛋白的系统发育分析表明,它们是新型的E1泛素激活同源物,是Helarchaeota E1-like和真核生物类泛素修饰物激活5蛋白的祖先。鉴于这种类型的宿主-病毒相互作用只在真核系统中被描述过,这很有趣。
阿斯加德病毒含有自主DNA甲基化和糖基化的基因,可能在宿主相互作用中发挥作用。DNA甲基化已被记录在噬菌体和Mimiviridae中,作为一种表观遗传修饰,用于逃避宿主的防御,如限制性修饰(RM)系统,劫持转录机器,调节复制周期,和/或防御具有RM系统的竞争病毒。有趣的是,Guaymas盆地的阿斯加德病毒可能能够自主地进行DNA甲基化,因为它们编码了5-甲基胞嘧啶、N4-甲基胞嘧啶和D12类N6-甲基腺嘌呤甲基转移酶。限制性内切酶(RE)类基因的同时出现,如Nidhogg中的RecB家族RE,可能表明功能性RM系统可以抑制宿主基因的表达,并将转录从宿主转移到病毒。Fenrir、Nidhogg和Ratatoskr也编码己糖转移酶,这是一类糖基转移酶(GTs),可能赋予修改DNA、蛋白质或脂质上的己糖的能力,以逃避宿主防御系统和抑制宿主分子功能。GTs已经在噬菌体和NCLDVs中被确认,但在古菌病毒中没有充分的记录。Fenrir编码两个重叠的基因,其序列与Sulfolobus islandicus rod-shaped virus 1 uncharacterized GT(SIRV1-GT)和alpha-1,2-mannosyltransferase(WbdA)同源。这表明Fenrir可以产生甘露糖糖链,这些糖链被编码的SAM依赖性甲基转移酶(WbdD)终止,而不利用宿主机器。此外,Fenrir编码L-malate GT(BshA),这是一个在Asgards中普遍存在的基因,被认为参与了细菌和古菌中低分子量硫醇(LMWT)的生物合成。Ratatoskr编码一个N-乙酰-α-D-葡糖胺基L-苹果酸脱氢酶1(BshB1)的同源物,这个基因也与LMWT的生物合成有关。最后,Nidhogg编码一个与SIRV1-GT相匹配的GT,以及一个可能形成葡聚糖链的糖原合成酶(GlgA)同源物(图4A)。
鉴于阿斯加德古菌与真核生物的关系,有趣的是,阿斯加德感染的病毒具有与真核生物NCLDV相似的复制和调节机制(PCNA、PolB、核糖核苷酸还原酶、DNA连接酶、DNMP激酶和类复制因子C AAA ATP酶)。然而,允许NCLDVs中转录和翻译解耦的mRNA加帽基因似乎并不存在于这些病毒中。此外,它们的许多特征与以前研究的古菌病毒不同。例如,完整的Nidhogg基因组编码了homeobox和PrimPol蛋白域,而这些蛋白域在古菌病毒中是缺乏的。它们还含有泛素蛋白修饰系统的基因,这些基因以前在古菌病毒中没有描述过,而在真核生物病毒中用于病毒的繁殖。它们编码的结构蛋白让人联想到Caudovirales中的那些蛋白,然而它们的大部分蛋白组成却与这一目不同。阿斯加德病毒似乎同时具有古菌和真核生物的病毒特征,这与它们的宿主的进化位置相一致。对与阿斯加德古菌有关的病毒的首次描述,推进了我们对病毒在阿斯加德古菌的生态和进化中的作用的理解。
参考文献
Rambo, I.M., Langwig, M.V., Leão, P. et al. Genomes of six viruses that infect Asgard archaea from deep-sea sediments. Nat Microbiol (2022). DOI: https://doi.org/10.1038/s41564-022-01150-8
作者简介

Brett J. Baker
Brett Baker是德克萨斯大学奥斯汀分校海洋科学系和综合生物学系的海洋微生物学副教授。他的研究旨在了解未培养的细菌和古细菌的生态学和进化。他的实验室主要利用计算方法来描述海洋沉积物的基因组多样性。近年来,他的团队对未培养的基因组的重建,使人们首次看到了生命之树上新分支的生理能力。Baker博士的研究小组还使用各种基于活动的方法来跟踪自然界中这些新型微生物的代谢活动。他的实验室在表征与真核生物有关的古菌方面也发挥了作用,这些古细菌推动了我们对复杂细胞生命起源的理解。2014年,他在密歇根大学获得了地质科学博士学位,在那里他研究了深海热液羽流和河口沉积物的地质微生物学。在此之前,他曾在加州大学伯克利分校担任过10年的研究助理,研究与酸性矿井排水有关的微生物。
猜你喜欢
iMeta简介 高引文章 高颜值绘图imageGP 网络分析iNAP
iMeta网页工具 代谢组MetOrigin 美吉云乳酸化预测DeepKla
iMeta综述 肠菌菌群 植物菌群 口腔菌群 蛋白质结构预测
10000+:菌群分析 宝宝与猫狗 梅毒狂想曲 提DNA发Nature
一文读懂:宏基因组 寄生虫益处 进化树 必备技能:提问 搜索 Endnote
生物科普: 肠道细菌 人体上的生命 生命大跃进 细胞暗战 人体奥秘
写在后面
为鼓励读者交流快速解决科研困难,我们建立了“宏基因组”讨论群,己有国内外6000+ 科研人员加入。请添加主编微信meta-genomics带你入群,务必备注“姓名-单位-研究方向-职称/年级”。高级职称请注明身份,另有海内外微生物PI群供大佬合作交流。技术问题寻求帮助,首先阅读《如何优雅的提问》学习解决问题思路,仍未解决群内讨论,问题不私聊,帮助同行。
点击阅读原文,跳转最新文章目录阅读
边栏推荐
- "In Vietnam, money is like lying on the street"
- Machine learning concept drift detection method (Apria)
- 大厂面试总结大全二
- Android uses sqliteopenhelper to flash back
- Weima, which is going to be listed, still can't give Baidu confidence
- An example of multi module collaboration based on NCF
- 项目通用环境使用说明
- Superscalar processor design yaoyongbin Chapter 7 register rename excerpt
- .NET ORM框架HiSql实战-第二章-使用Hisql实现菜单管理(增删改查)
- SIGMOD’22 HiEngine论文解读
猜你喜欢
Load test practice of pingcode performance test
VMware Tools和open-vm-tools的安装与使用:解决虚拟机不全屏和无法传输文件的问题

I wrote a learning and practice tutorial for beginners!
使用3DMAX制作一枚手雷
一直以为做报表只能用EXCEL和PPT,直到我看到了这套模板(附模板)
Machine learning concept drift detection method (Apria)
![[HCIA continuous update] WAN technology](/img/31/8e9ed888d22b15eda5ddcda9b8869b.png)
[HCIA continuous update] WAN technology

How to improve development quality
用于图数据库的开源 PostgreSQL 扩展 AGE被宣布为 Apache 软件基金会顶级项目

要上市的威马,依然给不了百度信心
随机推荐
力扣刷题日记/day5/2022.6.27
78岁华科教授冲击IPO,丰年资本有望斩获数十倍回报
. Net ORM framework hisql practice - Chapter 2 - using hisql to realize menu management (add, delete, modify and check)
How is the entered query SQL statement executed?
2022年全国CMMI认证补贴政策|昌旭咨询
Open source PostgreSQL extension age for graph database was announced as the top-level project of Apache Software Foundation
People in the workplace with a miserable expression
【系统盘转回U盘】记录系统盘转回U盘的操作
中断的顶半部和底半部介绍以及实现方式(tasklet 和 工作队列)
MySQL常用增删改查操作(CRUD)
力扣刷题日记/day1/2022.6.23
爬虫初级学习
android使用SQLiteOpenHelper闪退
Summary of subsidy policies across the country for dcmm certification in 2022
Why are some online concerts always weird?
力扣刷题日记/day6/6.28
大厂面试总结大全二
celebrate! Kelan sundb and Zhongchuang software complete the compatibility adaptation of seven products
【Hot100】31. 下一个排列
Superscalar processor design yaoyongbin Chapter 7 register rename excerpt